The Next Frontier of Genomic Medicine: Moving Beyond the CRISPR-Cas9 Revolution
The landscape of genome editing has been defined by the rapid ascent of CRISPR-Cas9, a tool that transformed molecular biology from a niche discipline into a high-octane engine of clinical innovation. Yet, even as CRISPR-Cas9 and its predecessors—zinc finger nucleases (ZFNs) and transcription activator-like effector nucleases (TALENs)—have matured, the field faces a stubborn reality: the "molecular scalpel" is not a universal solution.
While these tools are adept at correcting single-nucleotide point mutations, they often struggle with the complex architectural requirements of genetic medicine. Many debilitating diseases are caused by large-scale structural variations—deletions, duplications, inversions, and translocations—that require the precise insertion or rearrangement of entire gene-sized payloads. As scientists look toward the next generation of genetic cures, the "holy grail" has shifted from simply cutting DNA to sophisticated, large-scale genomic engineering.
A Chronology of Genomic Editing: From Cutting to Rewriting
The trajectory of genome editing can be viewed as a shift from destructive to constructive power. Early platforms like ZFNs and TALENs relied on protein-DNA recognition, which, while revolutionary, proved cumbersome to program. The advent of CRISPR-Cas9 in the early 2010s simplified targeting through RNA-DNA base pairing, but it remained fundamentally dependent on the cell’s own repair machinery, which is often error-prone and inefficient for large-scale insertions.
As researchers realized that the Double-Strand Break (DSB) approach—the hallmark of CRISPR-Cas9—was not always safe or effective for complex therapeutic needs, a new wave of innovation emerged. By the mid-2020s, the focus pivoted toward "DNA writing" tools: bridge recombinases, large serine recombinases (LSRs), and CRISPR-associated transposases (CASTs). These tools, coupled with advancements in immune-evasive DNA cargoes like circular single-stranded DNA (cssDNA), represent a paradigm shift: moving away from the "cut-and-pray" method toward a precise, programmable architecture.
The Bridge Recombinase: A New Class of RNA-Guided Precision
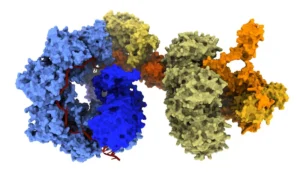
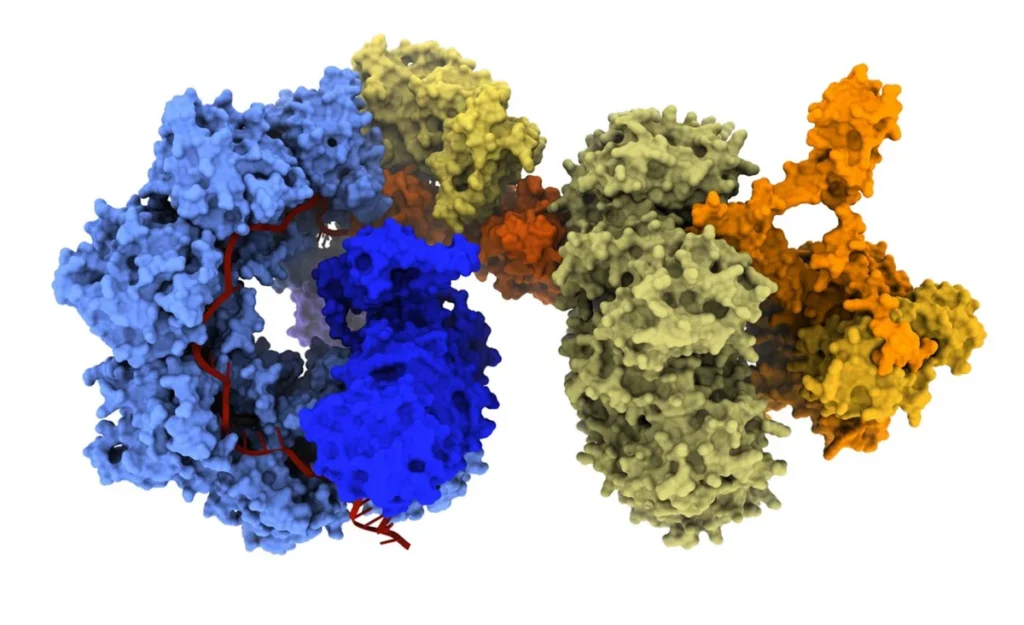
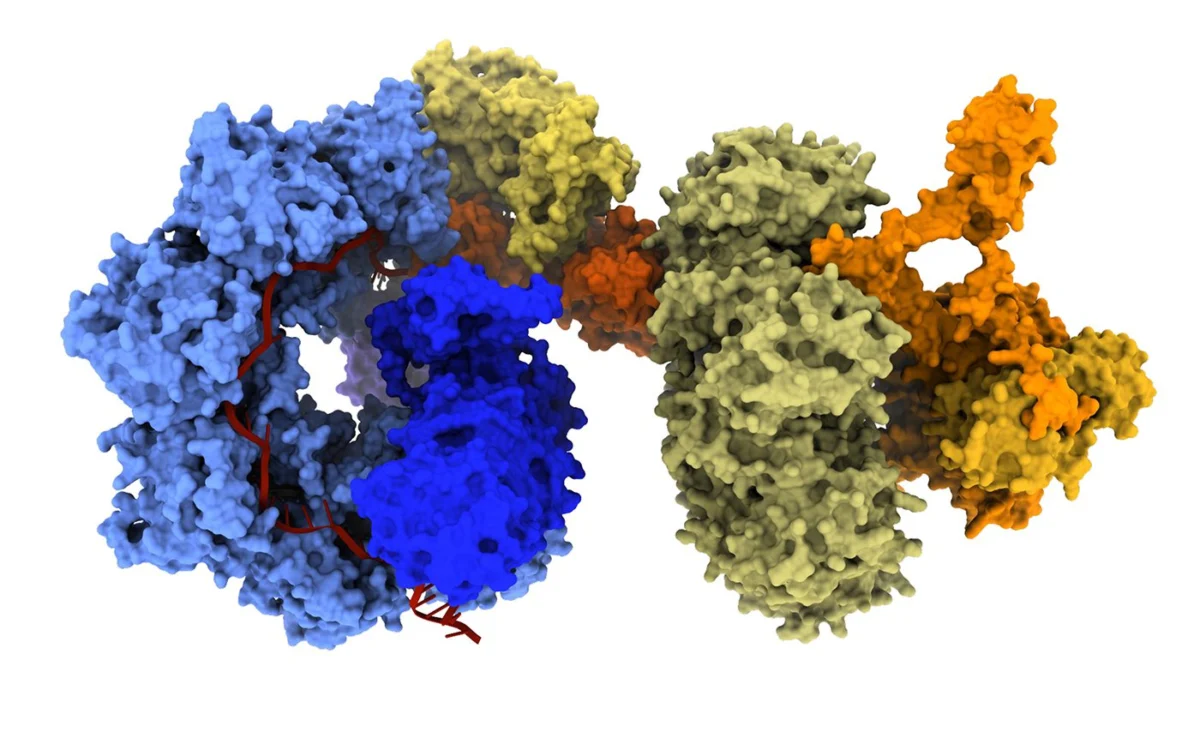
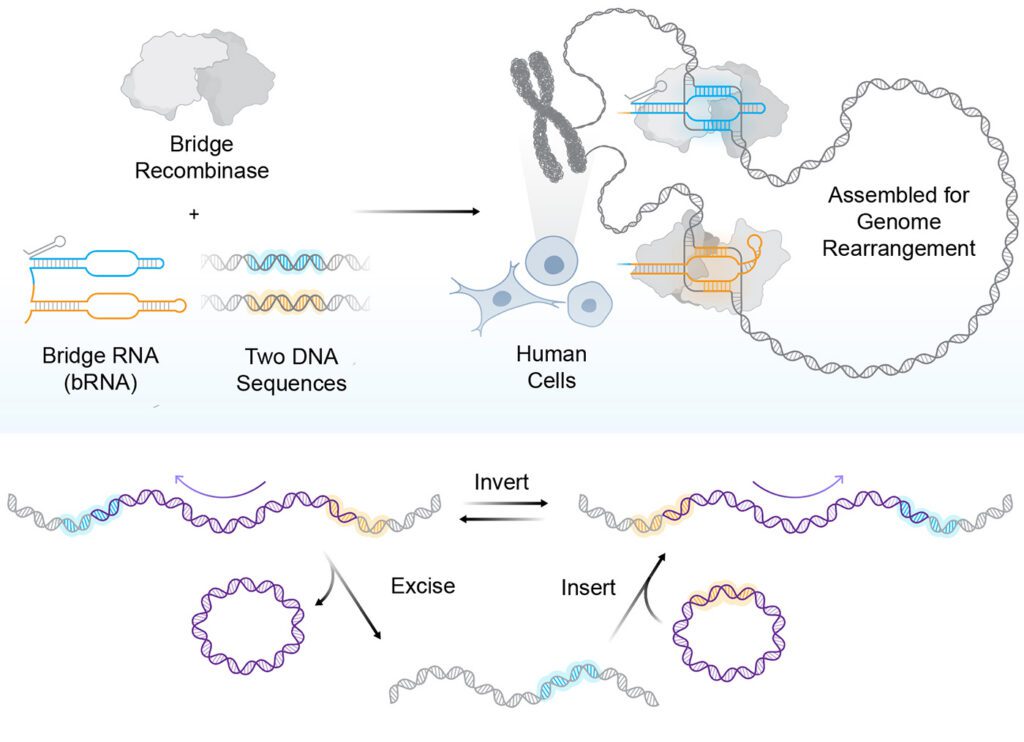
At the forefront of this evolution are bridge recombinases, the first truly programmable, RNA-guided DNA recombinases. Unlike traditional nucleases, bridge recombinases do not rely on creating harmful DSBs to initiate change. Instead, they function as a two-component system: a recombinase enzyme that catalyzes the DNA rearrangement and a bridge RNA guide that orchestrates the process.
The "bridge" itself is a masterpiece of modular engineering, featuring two independently programmable loops. One loop targets the specific genomic locus, while the other captures the donor DNA payload. By simply reprogramming the bridge RNA, researchers can dictate whether the system performs an insertion, an excision, or an in-place inversion.
"The most immediate advantage is the scale of DNA that bridge recombinases can manipulate," says Dr. Patrick Hsu, co-founder of the Arc Institute and an assistant professor of pathology at Stanford University. "A technology that can operate at the scale of whole-gene replacement or correct structural variants opens up a new class of genetic interventions."
Recent demonstrations have shown the system capable of multi-kilobase insertions and inversions of up to 0.93 Mb, a massive leap over the limited capacity of conventional CRISPR. Because these systems operate through a covalent intermediate rather than DSBs, they hold particular promise for post-mitotic cells—tissues that do not divide and are often recalcitrant to traditional gene-editing techniques.
Large Serine Recombinases: The High-Efficiency Standard
While bridge recombinases offer incredible flexibility, other applications demand extreme precision at fixed "safe harbor" genomic sites. For these scenarios, researchers are turning to large serine recombinases (LSRs).
LSRs are prized for their unidirectional nature, which ensures that once a large DNA payload is inserted into the genome, it stays there with high stability. In 2025, a landmark study published in Nature Biotechnology detailed an engineered LSR capable of multi-kilobase insertions with 53% efficiency and 97% genome-wide specificity.
The implications for clinical medicine are profound. Stylus Medicine, a company co-founded by Hsu, is currently advancing these LSRs for in vivo therapies. The ability to utilize LSRs in non-dividing cells, such as primary human T cells, creates an immediate pathway for ex vivo cell therapies in oncology and autoimmune disease management.
CASTs and the Power of Directed Evolution
CRISPR-associated transposases (CASTs) represent another sophisticated branch of this tree. By utilizing nuclease-deficient CRISPR machinery, CASTs guide site-specific DNA transposition without the need for DSBs.
Early iterations of CASTs faced a significant hurdle: their activity in human cells was negligible. This changed through the use of PACE (phage-assisted continuous evolution), a platform developed by the lab of Dr. David Liu at the Broad Institute of MIT and Harvard. By linking the desired integration activity to the replication of bacteriophages, the researchers were able to force the rapid evolution of the CAST protein components.
"In PACE, you link the desired activity of the evolving biomolecule to the expression of this essential gene," explains Dr. Isaac Witte of Harvard University. This process led to the creation of "evoCAST," an optimized system that provides over 200-fold improved integration activity. With the ability to mobilize payloads ranging from 1 kb to 15 kb, evoCAST offers a versatile toolkit for human genome writing.
Solving the Payload Problem: The Promise of cssDNA
Even with the most precise editing tools, the bottleneck remains delivery. Most viral and non-viral vectors are size-limited, and the innate toxicity of foreign double-stranded DNA (dsDNA) often triggers an immune response in the host.
Full Circle Therapeutics has pioneered a solution with "C4DNA"—a circular, clean, concealed, and customizable single-stranded DNA (cssDNA) format. "The holy grail of gene editing is kilobase DNA integration," notes Dr. Howard Wu, co-founder and CSO of Full Circle Therapeutics. "We address the challenge from the donor side."
Through serendipitous discovery and rigorous benchmarking, the team found that cssDNA outperformed linear DNA in integration tasks. By developing technologies like INSTALL (Integration through Nucleus-synthesized Template Addition of Large Lengths), they have created a platform that works in tandem with various nucleases and recombinases to achieve high-fidelity gene-sized writing while remaining immune-silent.
Supporting Data and Technical Challenges
Despite the excitement, the field remains cautious regarding the current metrics.
| Technology | Primary Mechanism | Key Advantage | Current Challenge |
|---|---|---|---|
| Bridge Recombinase | RNA-guided rearrangement | Modular, non-DSB | Efficiency (approx. 20%) |
| LSRs | Site-specific integration | High stability/specificity | Limited RNA-programmability |
| evoCAST | Site-specific transposition | Large payloads (up to 15kb) | Molecular complexity |
| cssDNA (C4DNA) | Immune-evasive template | Low immunogenicity | Delivery size constraints |
Dr. Hsu acknowledges that while current proof-of-concept numbers are promising, the next phase of development must focus on safety and efficiency. "Improving both metrics will be necessary for safe and effective therapeutic applications," he notes. Similarly, Dr. Witte emphasizes that the molecular complexity of tools like evoCAST—which requires seven distinct protein subunits—must be streamlined to facilitate clinical translation.
Implications for the Future of Medicine
The shift toward these "writer" technologies changes the fundamental calculus of gene therapy. We are no longer looking at gene editing as a way to "fix" a spelling error; we are looking at it as a way to "install" functional software into the human genome.
The clinical implications are vast. For diseases involving large structural defects, such as certain muscular dystrophies or rare metabolic disorders, the ability to insert a functional gene copy into a safe harbor site—or to excise a deleterious mutation without risking genomic instability—could be curative.
As the industry moves from basic discovery toward GMP-grade production and clinical trials, the collaborative nature of this field is striking. Whether it is through the refinement of recombinases or the development of immune-stealth DNA, the goal remains the same: creating a "one-and-done" treatment that is both safe and permanent.
The quest for the holy grail of genome editing is far from over, but the tools currently being forged in the lab promise a future where "deplorable diseases" are not just managed, but systematically rewritten. The next decade of genomic medicine will likely be defined not by the speed at which we can cut, but by the precision with which we can write.